
























Nori Human L-Selectin ELISA Kit
$461.00 – $832.00
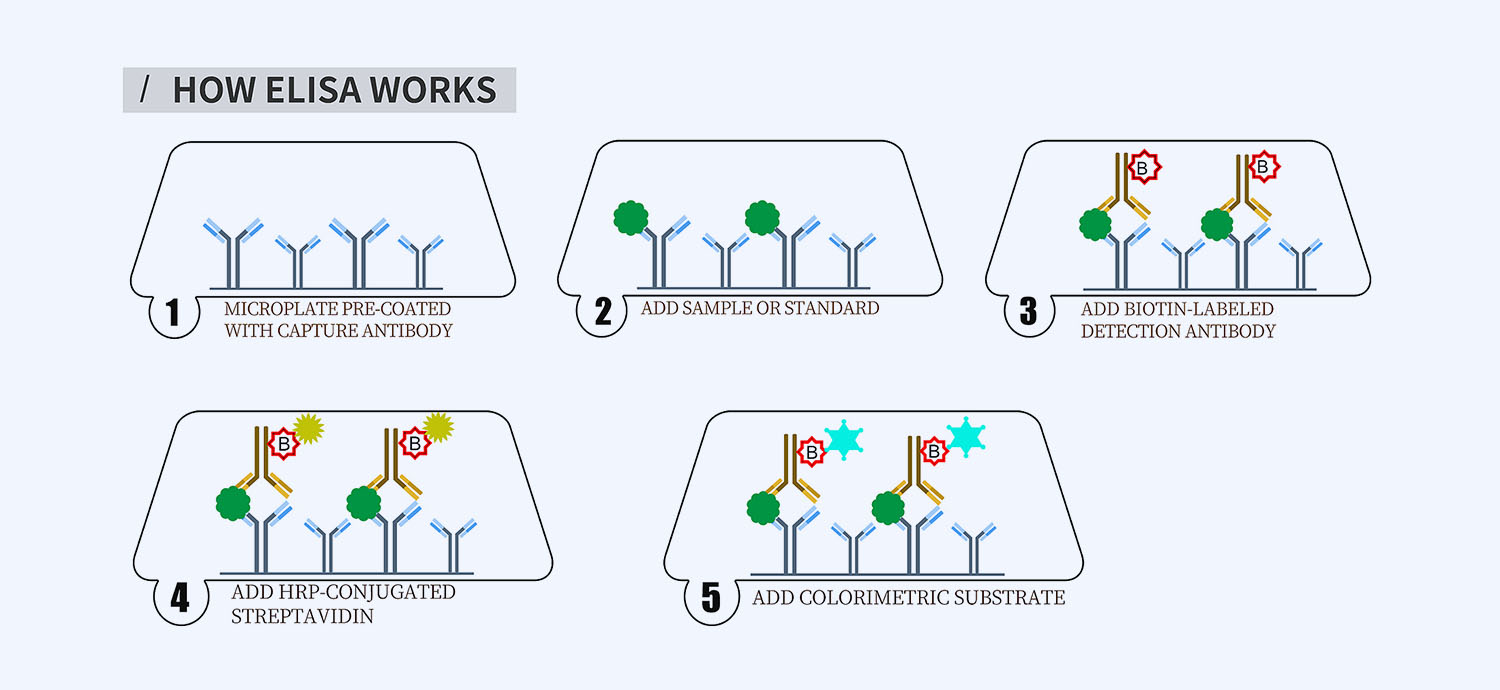
This ELISA kit is for quantification of L-Selectin in human. This is a quick ELISA assay that reduces time to 50% compared to the conventional method, and the entire assay only takes 3 hours. This assay employs the quantitative sandwich enzyme immunoassay technique and uses biotin-streptavidin chemistry to improve the performance of the assays. An antibody specific for SELL has been pre-coated onto a microplate. Standards and samples are pipetted into the wells and any SELL present is bound by the immobilized antibody. After washing away any unbound substances, a detection antibody specific for SELL is added to the wells. Following wash to remove any unbound antibody reagent, a detection reagent is added. After intensive wash a substrate solution is added to the wells and color develops in proportion to the amount of SELL bound in the initial step. The color development is stopped, and the intensity of the color is measured.
Alternative names for L-Selectin: L-selectin (SELL), CD62L
This product is for laboratory research use only not for diagnostic and therapeutic purposes or any other purposes.
- Description
- How Elisa Works
- Product Citation
- Reviews (0)
Description
Nori Human L-Selectin ELISA Kit Summary
Alternative names for L-Selectin: L-selectin (SELL), CD62L
| Assay Type | Solid Phase Sandwich ELISA |
| Format | 96-well Microplate or 96-Well Strip Microplate |
| Method of Detection | Colorimetric |
| Number of Targets Detected | 1 |
| Target Antigen Accession Number |
P14151 |
| Assay Length | 3 hours |
| Quantitative/Semiquantitative | Quantitative |
| Sample Type | Plasma, Serum, Cell Culture, Urine, Cell/Tissue Lysates, Synovial Fluid, BAL, |
| Recommended Sample Dilution (Plasma/Serum) | No dilution for sample <ULOQ; sufficient dilution for samples >ULOQ |
| Sensitivity | 60 pg/mL |
| Detection Range | 0.312-20 ng/mL |
| Specificity | Natural and recombinant human L-Selectin |
| Cross-Reactivity | < 0.5% cross-reactivity observed with available related molecules, < 50% cross-species reactivity observed with species tested. |
| Interference | No significant interference observed with available related molecules |
| Storage/Stability | 4 ºC for up to 6 months |
| Usage | For Laboratory Research Use Only. Not for diagnostic or therapeutic use. |
| Additional Notes | The kit allows for use in multiple experiments. |
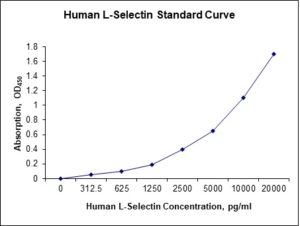
Standard Curve
Kit Components
1. Pre-coated 96-well Microplate
2. Biotinylated Detection Antibody
3. Streptavidin-HRP Conjugate
4. Lyophilized Standards
5. TMB One-Step Substrate
6. Stop Solution
7. 20 x PBS
8. Assay Buffer
Other Materials Required but not Provided:
1. Microplate Reader capable of measuring absorption at 450 nm
2. Log-log graph paper or computer and software for ELISA data analysis
3. Precision pipettes (1-1000 µl)
4. Multi-channel pipettes (300 µl)
5. Distilled or deionized water
Protocol Outline
1. Prepare all reagents, samples and standards as instructed in the datasheet.
2. Add 100 µl of Standard or samples to each well and incubate 1 h at RT.
3. Add 100 µl of Working Detection Antibody to each well and incubate 1 h at RT.
4. Add 100 µl of Working Streptavidin-HRP to each well and incubate 20 min at RT.
5. Add 100 µl of Substrate to each well and incubate 5-30 min at RT.
6. Add 50 µl of Stop Solution to each well and read at 450 nm immediately.
Background:
L-selectin (SELL), also known as CD62L, is a cell adhesion molecule found on leukocytes and the preimplantation embryo. It belongs to the selectin family of proteins, which recognize sialylated carbohydrate groups.[1] It is cleaved by ADAM17. SELL is a cell surface component that is a member of a family of adhesion/homing receptors that play important roles in lymphocyte-endothelial cell interactions. L-Selectin is composed of multiple domains: one homologous to lectins, one to epidermal growth factor, and two to the consensus repeat units found in C3/C4-binding proteins. L-selectin acts as a “homing receptor” for lymphocytes to enter secondary lymphoid tissues via high endothelial venules. Ligands present on endothelial cells will bind to lymphocytes expressing L-selectin, slowing lymphocyte trafficking through the blood, and facilitating entry into a secondary lymphoid organ at that point. Central memory T-lymphocytes, which have encountered antigen, express L-selectin to localize in secondary lymphoid organs. High expression of L-selectin on human bone marrow progenitor cells is an early sign of cells becoming committed to lymphoid differentiation.[2] L-selectin acts as a receptor to facilitate adhesion of the embryo to the site of invasion on the surface epithelium of the uterine endometrium. Removal of MUC-1 exposes the oligosaccharide ligands of the uterine epithelium, thus allowing binding by the L-selectin receptor of the trophopblast cell, followed by embryo adhesion and invasion. L-selectin activates the Ras pathway via the tyrosine kinase p56lck.[3] L-selectin from human, but not from mouse neutrophils binds directly to E-selectin.[4] Podocalyxin-like protein and human complement Factor H are ligands for L-selectin.[5][6] Nucleolin expressed on the surface of head and neck squamous cell carcinoma cells is an active ligand for L-selectin.[7] Reduced CD62L expression suggests blood neutrophils have undergone priming in COPD but not in asthma. The increased shedding of CD62L receptor by COPD blood neutrophils suggests a high sensitivity for activation.[8] CD62L is a functional and phenotypic marker for circulating innate lymphoid cell precursors.[9] L-selectin has been identified as an HIV-1 adhesion receptor on CD4+ T cells.[10]
References
- Prakobphol A, et al. (1998). Biochemistry. 37 (14): 4916–27.
- Kohn LA, et al. (2012). Nat. Immunol. 13 (10): 963–71.
- Brenner B, et al. (1997). Proc. Natl. Acad. Sci. U.S.A. 93 (26): 15376–81.
- Zöllner O, et al. (1997). J. Cell Biol. 136 (3): 707–16.
- Sassetti C, et al. (1998). J. Exp. Med. 187 (12): 1965–75.
- Malhotra R, et al. (1999). Biochem. J. 341 (1): 61–9.
- Goldson TM, et al. (2020) Am. J. Physiol., Cell Physiol. 318 (1), C83-C93.
- Lokwani R, et al. (2019) Int J Chron Obstruct Pulmon Dis 14, 2517-2525.
9. Bar-Ephraim YE, et al. (2019) J. Immunol. 202 (1), 171-182.
- Kononchik J, et al. (2018) Nat Commun 9 (1), 2825.

Reviews
There are no reviews yet.